¿Para qué queremos tantos escarabajos diferentes?


La evolución conduce hacia un número cada vez mayor de especies, y eso nos complica mucho conocer cuántas habitan la Tierra. ¿Hay que destinar esfuerzos a conservarlas o con unas pocas ya tendríamos suficiente?
![]()
Para Jordi y Olga.
En 1992 Edward O. Wilson popularizó el concepto de 'biodiversidad' gracias su fantástico libro The Diversity of Life. Desde entonces, la conservación de las especies se ha incorporado a los valores de nuestra sociedad. Pero ¿cuál es la magnitud de ese esfuerzo? Saber cuál es el número de especies que habitan el planeta puede ayudarnos a responder la pregunta. Nuestra sociedad se siente atraída los números, y cuanto más extremos son, mayor atracción despiertan. En 2015 tuvo una gran repercusión un artículo de T. W. Crowther y colaboradores publicado en la revista Nature en el que cartografiaban la densidad de árboles en la Tierra y estimaban en 3,04 · 1012 el número de árboles vivos en todo el planeta. Rápidamente hubo que calcular el número de árboles de nuestros bosques —obteniendo su correspondiente cuota de popularidad—. Para los interesados en los números diremos que ese valor era aproximadamente de 7 · 109 para los bosques de España.
Pero esas estimas tienen sus limitaciones, ya que se basan en una serie de asunciones que no se cumplen siempre. Jonathan Chase presentó no hace mucho tiempo una ponencia en un congreso de biogeografía en la que debatía los problemas de las extrapolaciones que calculan el número de especies que viven en la Tierra. Estas extrapolaciones tienen mucha incerteza y se han dado valores que oscilan entre los 3 y los 100 millones de especies —uno de los cálculos daba en 2011 una cifra de 8,7 millones de eucariotas—.
Para obtener estos números un método habitual es extrapolar a áreas mayores el número de especies —lo que denominamos riqueza— identificadas en una zona de extensión conocida. Así en última instancia se puede estimar la riqueza de especies de todo el planeta. Este método permite tener una idea del número de especies que aún no se han identificado, básicamente porque no se ha prospectado los sitios en los que viven. Esto tiene implicaciones para la conservación, porque si el hábitat que existe en ese sitio se pierde, esas especies también.
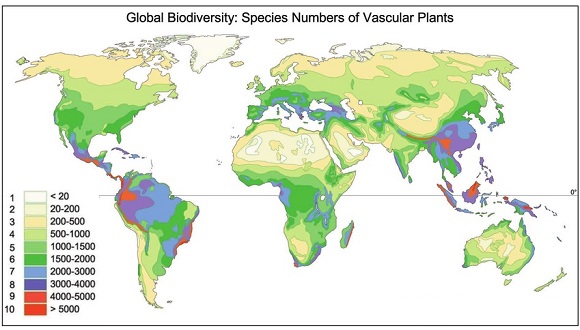
Fuente: Barthlott, Kier, Rafiqpoor, Kreft, Krüper & Mutke. 2004. Nees Institute for Biodiversity of Plants, University of Bonn. W. Barthlott 1996, 2004. En el artículo: Mutke J., Barthlott W. (2005). Patterns of vascular plant diversity at continental to global scale. Biol. Skr. 55:521-531. ISSN: 0366-3612. ISBN 87-7304-304-4.
Lógicamente hay que hacer correcciones porque esas extrapolaciones no son iguales para todos los grupos de seres vivos. Los hay con muchas especies endémicas —como los coleópteros— y por tanto prospectar nuevas áreas hace aumentar rápidamente su riqueza de especies. Otros grupos —como los musgos— se encuentran ampliamente distribuidos y aumentar el área no incrementa demasiado su número. Por otro lado, la riqueza de especies no es la misma en todos los hábitats. Por tanto, la extrapolación del número de especies a superficies mayores debería analizar por separado la relación entre el número de especies de cada grupo y el aumento del área de cada uno de los hábitats, y luego ponderarlo por la abundancia de esos hábitats. También hay que valorar cómo afecta a estas extrapolaciones el hecho de que los hábitats estén más o menos conectados en paisajes fragmentados.
Existe además otra consideración importante: el aumento de la riqueza de especies en áreas mayores no es lineal. Sigue una curva con una asíntota que indica que el ambiente tiende a saturarse de especies. Aunque prospectemos con gran intensidad, cuesta mucho encontrar nuevas especies, como saben los naturalistas que buscan novedades. La razón es que el número de especies de la región que son capaces de vivir en un hábitat es limitado ya que se tienen que repartir los recursos entre ellas. Esta regla es bien conocida y se explica en las primeras clases de los cursos de ecología general. Es bastante curioso que en un congreso especializado el ponente tenga que recordar a los oyentes principios básicos de ecología, fundamentándose en modelos matemáticos más o menos sofisticados y en grandes bases de datos. Supongo que indica de que hay principios suficientemente robustos como para que persistan conforme progresa la ciencia, aunque el deslumbramiento que provocan los nuevos métodos los condene momentáneamente al olvido.
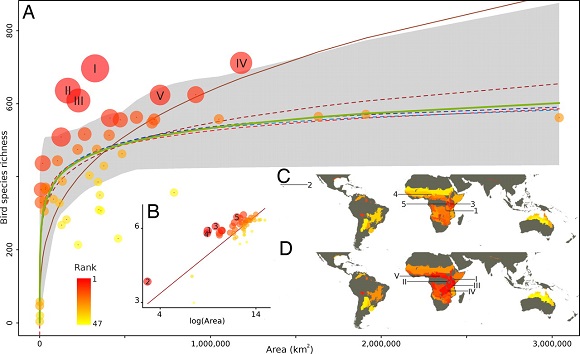
Fuente: Guilhaumon F., Gimenez O., Gaston KJ., Mouillor D. (2008). Taxonomic and regional uncertainty in species-area relationships and the identification of richness hotspots. PNAS 105 (40): 15458-15463. DOI: https://doi.org/10.1073/pnas.0803610105
Una de las conclusiones de Jonathan Chase es que estamos sobrevalorando el número de especies que viven en la Tierra. No creo que eso nos tenga que preocupar excesivamente, a no ser que estemos realmente interesados en que dicho número conste en el libro Guinness de records. El número de especies que habita en el planeta sigue siendo el más elevado de la historia de la Tierra. Es el resultado de una evolución que tiende a generar nuevos linajes siempre que sean compatibles —a veces incluso con un alto grado de optimización— con la gran heterogeneidad ambiental existente. A su vez, la propia diversidad de la biota tiende a acrecentar la heterogeneidad ambiental, generando nuevos hábitats o produciendo nuevos recursos. Uno de los hechos más notables que se ha producido en la ciencia durante las últimas décadas de la naturaleza es el reconocimiento de un desmesurado número de genotipos que atribuimos a especies diferentes a partir de su DNA recolectado en el medio. Estos métodos moleculares se han aplicado a los microorganismos, y se han descubierto multitud de especies, identificadas exclusivamente por su dotación genética - técnicamente denominadas OTUs —Operational Taxonomic Units—. Aunque estos métodos no están exentos de dudas, en general se acepta que el número de especies de microorganismos es más elevado de lo que se esperaba. A modo de ejemplo, se ha estimado que en un litro de agua de mar puede haber 20.000 de esas "especies moleculares" e incluso se han encontrado cientos de ellas en rocas a profundidades de 1.000 metros o más. Esto hace que actualmente haya estimaciones de la biodiversidad global de la Tierra que alcanzan valores de entre 1·109 y 6·109 especies.
De todas formas, la conservación de la biodiversidad tiene suficientemente trabajo incluso con las especies que conocemos. Aunque el número de las especies desconocidas no sea en realidad tan enorme como el que dicen las estimaciones, no hay duda de que nuestro conocimiento de la biodiversidad es aún incompleto. Debemos trabajar con esa premisa y con la certeza de que el tiempo corre en contra. El desconocimiento del detalle de la biología de muchas especies —de las desconocidas, pero también de muchas de las descritas por los científicos naturalistas—, junto a las evidencias de que la biodiversidad tiene un papel importante en la regulación del funcionamiento de los ecosistemas ha promovido el éxito del concepto de especie funcional —o tipo de especies funcionales—. Se trata de agrupar las especies por aquellas de sus características que tienen una implicación similar en el funcionamiento del ecosistema. Esta aproximación ha tenido un éxito notable entre investigadores que pretenden cuantificar el funcionamiento de la biosfera a escala global. Por ejemplo, cuando se calculan balances de carbono, es esencial determinar la producción primaria de los ecosistemas, la cual básicamente se debe a la asimilación fotosintética del CO2 atmosférico por la vegetación. Desde este punto de vista, todas las especies que fotosintetizan se podrían poner juntas en un gran grupo funcional.
Obviamente esta aproximación puede causar horror a un naturalista enamorado de la biodiversidad y de las particularidades de las respuestas evolutivas. Pero para alguien que quiere medir la producción primaria de todos los bosques del mundo, no es tan importante si quien realiza la fotosíntesis es una conífera o una fagácea. En realidad, los modelos que describen el funcionamiento de los ecosistemas a escala global han progresado y están en condiciones de considerar subgrupos funcionales con comportamiento fisiológico diferenciado.

Fuente: Friends of the Earth International (CC BY 2.0)
Pronto surgió la duda sobre la importancia de la diversidad de especies a la hora de determinar los grandes flujos de los ecosistemas, como la producción primaria. Si diferentes especies hacen funciones similares, con unas pocas sería suficiente para que los ecosistemas funcionaran. Es lo que se llamó redundancia funcional. Además, es bastante probable que las especies responsables de un determinado flujo de energía o materia sean particularmente abundantes. Por ejemplo, en cultivos que necesitan de polinizadores, para obtener una buena cosecha sólo se necesita un número reducido de especies dominantes de insectos. Este planteamiento puso de los nervios a quienes luchaban por preservar la biodiversidad del planeta. ¿Vale la pena todo el esfuerzo de conservar tal ingente variedad de especies cuando a efectos prácticos sirven para lo mismo, suponiendo que realmente sirvan para algo? Obviamente el planteamiento contiene muchas simplificaciones, pero no hay que tomárselo a la ligera. La primera simplificación es asumir que existe una única función del ecosistema que merezca ser preservada. Obviamente no es así, y las especies redundantes para una función no tienen por que serlo para otra función. Además, el nivel de redundancia de especies no tiene por que ser igual para todas las funciones. Existen algunas funciones que son realizadas por pocas especies, como es el caso de la fijación del nitrógeno inerte de la atmósfera en formas asimilables, en las que intervienen un número reducido de vegetales en connivencia con unos cuantos microorganismos. Si desaparecen, las consecuencias son desproporcionadas. Son las llamadas especies clave.
Esta cuestión se ha convertido en una de las líneas de investigación más atrayentes de las últimas décadas: conocer la contribución de la biodiversidad en el funcionamiento de los ecosistemas. Se partía de planteamientos teóricos que abogaban por una relación positiva entre biodiversidad y funcionamiento de los ecosistemas. El razonamiento se basaba en la complementariedad de las especies: lo que no sabía hacer una especie, lo podía hacer otra. El propio Edward O. Wilson —junto a su colega Robert MacArthur, con quien desarrolló unos sencillos y exitosos modelos matemáticos para explicar la riqueza de especies en medios insulares— había apoyado esta idea. Paul Ehrlich y su esposa Anne introdujeron además la inquietante idea de que la redundancia tenía un límite impreciso, como los tornillos que aguantan el fuselaje de un avión: podemos perder alguna pieza y el avión sigue volando, hasta que la caída de un único tornillo adicional puede conducir de golpe y en cualquier momento a la catástrofe. Tampoco se descartaba un comportamiento idiosincrático de cada ecosistema, con las consiguientes dificultades para establecer unas reglas generales que ayudaran a establecer estrategias de conservación a priori. No obstante, la existencia de redundancia funcional entre especies estaba ahí.
Cualquier ecólogo crítico se pregunta que relevancia tiene para el funcionamiento de un ecosistema, por no decir de la biosfera en su conjunto, que una mariquita tenga siete u ocho puntos negros sobre el fondo rojo de su caparazón. Se han realizado múltiples experimentos manipulando el número y el tipo de especies en condiciones de campo, invernadero o de laboratorio —los llamados mesocosmos— y midiendo las propiedades funcionales del ecosistema. Se han analizado grandes bases de datos de campo, como los inventarios forestales, con información sobre biodiversidad y medidas de producción primaria. Se han desarrollado modelos teóricos y simulaciones numéricas. Aunque quedan muchos detalles por averiguar sobre los mecanismos implicados, existe un consenso en que la contribución de la biodiversidad al funcionamiento de los ecosistemas se satura a partir de cierto número de especies. Es decir, la hipótesis de la relación positiva y la de la redundancia no son excluyentes y ambas se cumplen en cierta medida.

Autor: Francisco Lloret
En realidad, la redundancia no es un problema para la conservación de la biodiversidad, sino todo lo contrario. Vemos que la redundancia permite mantener el funcionamiento de los ecosistemas, una de cuyas principales cualidades es la regulación de los flujos de materia y energía a lo largo del tiempo, ya que las condiciones del medio cambian tanto en el espacio como en el tiempo. La redundancia de la biodiversidad proporciona un seguro ante un medio inexorablemente cambiante. Una especie que es redundante en un momento puede convertirse en clave bajo unas condiciones ambientales diferentes. Esas nuevas condiciones pueden ser perjudiciales para otras especies que antes realizaban funciones similares e incluso podían haber sido dominantes. De forma parecida, una especie dominante en un lugar, y por tanto responsable en gran medida del funcionamiento del ecosistema, puede dejar de serlo en otra localidad próxima. Es lo que se ha visto en un estudio reciente sobre el papel de los polinizadores en la producción de determinadas cosechas en la costa este de Norteamérica. Aunque en una localidad, unas pocas especies se bastan para proporcionar suficiente polen para obtener una buena cosecha, cuando nos desplazamos a lo largo del territorio se necesitan nuevas especies que cubran esa función. Si a alguien le queda duda de como redundancia y especies clave se complementan para proporcionar estabilidad a los ecosistemas, le invito a leer el breve texto que prepararon Paul Ehrlich y Brian Walker en el año 1998.
Volvemos pues a los modelos clásicos que interpretaban la esencia de la vida a través de su diversidad, ya que reconocían el papel de combinar abundancia y rareza. Las especies abundantes, con éxito, representan la optimización de soluciones biológicas eficientes frente a un medio concreto. Pero cuando el medio cambia, y siempre lo acaba haciendo, las especies raras tienen su oportunidad. Es fácil ver que este mismo razonamiento se aplica al papel de la diversidad en las sociedades humanas. Ramon Margalef nos explicaba que el interés de la biodiversidad reside en esas especies raras, latentes, que aparentemente juegan un papel escaso en el ecosistema, pero que son el potencial biológico que proporcionará soluciones en entornos futuros, diferentes. Vemos pues como seguimos dando vueltas alrededor de algunos principios robustos —fácilmente comprensibles para todos los humanos—, mientras la ciencia los matiza, acumula evidencias y así amplía nuestra capacidad de comprenderlos.


