Plant’s Lego


How are plants organized in the world? Why have they evolved this way? Francisco Lloret answers that after researcher Carlos Herrera was granted with the Haeckel Price in ecology.
![]()
To Carlos
In the recent ecology congress co-organized by the EEF (European Ecological Federation) and the BES (British Ecological Society), Carlos Herrera was granted with the Haeckel Prize, the highest European award in ecological research (Jaume Terradas mentioned this award in his post a few months ago). Carlos has been studying the mutualistic interactions between plants and animals for many years, that is, those processes, such as dispersion or pollination, that bring benefits to both parties. In addition to a great analytical capacity, Carlos is a great naturalist, with an extraordinary capacity for observation. We should not be surprised that, in his eagerness to clear new paths of knowledge, he spoke to us about the ecological significance of the variability among the organs (leaves, flowers, fruits) of the same plant. In this article you can find your complete presentation. I will limit myself to commenting on some ideas.
The reason for the success of the modules lies in their ability to adjust to small variations in the environment that represent opportunities to obtain resources or to support stress pulses.
Unlike most animals, plants are organized on the basis of modules —however, there are some animals, such as corals, which are also modular while they attached to a solid surface that attach to a substrate. Moduls are functional growth units, such as leaves, which are repeated in the same organism and have a certain degree of functional autonomy. In a vertebrate, the loss of most parts of the body, an arm or the head, is irreplaceable while in a plant the fine roots that absorb water and nutrients quickly perish and are replaced immediately. The reason for the success of the modules lies in their ability to adjust to small variations in the environment that represent opportunities to obtain resources or to support stress pulses.
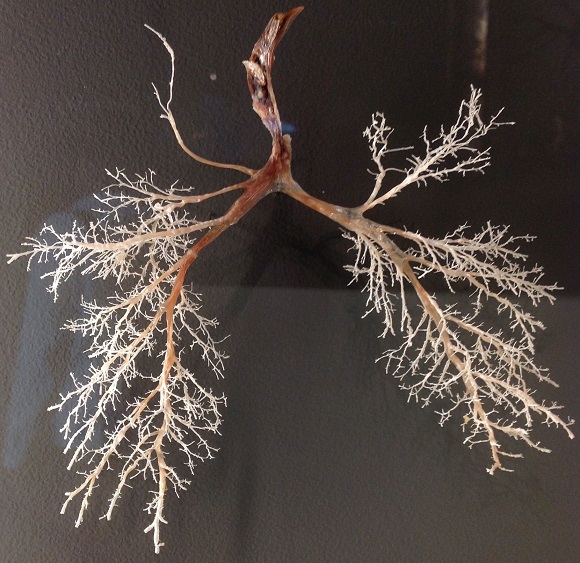
If life exists thanks to its ability to fix the energy of its surroundings, mainly from the radiation that comes from the Sun, why has not been organized into a single thick photosynthetic sheet that covers the entire Earth and completely captures radiation?
If life exists thanks to its ability to fix the energy of its surroundings, mainly from the radiation that comes from the Sun, why has not been organized into a single thick photosynthetic sheet that covers the entire Earth and completely captures radiation? The answer is that the environment is not homogeneous or constant. In a forest, the light arrives with different inclination throughout the day and the modules, as a whole, allow to better capture the variation in the arrival of the radiation. Similarly, the vast majority of plants prefer to distribute their reproductive effort in distinct units, the flowers, which can capture different sources of pollen, rather than concentrating it in a single flower. This explains the relationship between variability of different parameters of leaves growing in the same plant -such as the density and size of stomata- and plant fertility, as Carlos showed. It is perfectly consistent with an evolutionary interpretation of the modules, since its variation within an individual allows a better adjust to a variable environment.
Thus, an important source of intra-individual variability in plants can be attributed to the environment’s own variability. It is well known that the leaves of oak trees that grow exposed to the sun in the upper part of the crown are thicker and smaller than those that grow in the shade in the inner branches. But there is also an internal component of the plant that governs that variability. There are evolutionary lineages with less variable modules, as some studies have shown. The genetic legacy also manifests itself because there are lineages that develop certain forms of modular growth that are intrinsically less variable for anatomical or simply geometric reasons.
An important source of intra-individual variability in plants can be attributed to the environment’s own variability. It is well known that the leaves of oak trees that grow exposed to the sun in the upper part of the crown are thicker and smaller than those that grow in the shade in the inner branches.
For example, linear leaves —defined mostly by a single dimension— as those of grasses are probably less likely to be variable than the wider leaves of oaks —extended in a plane defined by two dimensions. Linear sheets, however, solve this disadvantage to a certain extent by easily folding, as Carlos Herrera recalls. These physical constrictions are very important in biology, although we often neglect them when searching for more teleological or finalist explanations. That is, not all biological forms obey an optimization criterion, instead they are often the result of opportunity. Indeed, evolution is a master of opportunity and works at ease with preexisting structures.

In turn, the variability of the organs within the same plant generates microenvironments that are used by other organisms. Molecular techniques of massive sequencing are allowing to recognize the diversity of microorganisms that inhabit the leaves surface, which has come to be called the ‘phyllosphere’. Surely the most interesting thing is that there are signs that this phyllosphere in turn can condition the functioning of the leaves and by extension of the whole tree and the ecosystem. For example, certain combinations of microorganisms could control infections that are caused by pathogens that penetrate through the stomata of the leaves. It is something similar to what happens in our intestine with its bacterial flora, as has been recently revealed, determining important physiological processes beyond the digestive function.
There are signs that this phyllosphere in turn can condition the functioning of the leaves and by extension of the whole tree and the ecosystem.
Going back to the plants, the roots are also highly variable modules growing within the huge heterogeneity of soils, at the level of centimeters or millimeters. It has been demonstrated that the roots of different species determine characteristic microbial communities since they generate different microenvironments, for example, through the different chemical composition of their exudates. On the other hand, soil microbial communities favor or limit the establishment and growth of different plant species, determining the interactions they establish between them. Thus, we see how the modularity of a plant, and its variability, affects other organisms, which, in turn, condition the functioning of the plant.

We face a feedback loop that deserves to be explored in depth as technical advances permit. For example, the epigenetic processes of regulation of gene expression in different cells or tissues of the same organism are in turn determined by environmental processes, and can constitute a mechanism through which these feedbacks are articulated. We often think on the interactions between different organisms forgetting these mutual feedback loops. It is easy to understand that a type of plant determines the microbial communities that live in its environment. It is also easy to think that soil microbial communities can provide different soil nutrients in a way that may favor certain plants. What is not so obvious is that the same type of plant can be favored or harmed by itself or by another type of plant through its effects on third parties, and that these effects are transferred over time, even when the former plants disappear from a given site.
If the intraindividual variability is so important, does that mean that we are forced to measure all the modules of all the plants in order to understand how a forest, a shrobland or a grassland works?
Finally, if the intraindividual variability is so important, does that mean that we are forced to measure all the modules of all the plants in order to understand how a forest, a shrobland or a grassland works? It is the typical problem of scaling between levels of study. It is equivalent to asking ourselves if we have to measure all the trees in order to explain a forest landscape. The basic rules of scaling tell us that when we add data from different units (leaves of a plant, trees of a forest) the averages are good indicators of the whole. While when we disaggregate the information from broad study units, we need measures of the variability within these units to be able to distinguish at some extent between subunits. But if the environment is very changing along time, the spatial variability between the different units or modules is fundamental to explain properties of the system related to its stability or dynamics. Thus, the measure of the intraindividual variability of the leaves of a tree is important to understand the overall efficiency of the whole tree in front of the Sun’s journey throughout the sky along the day or the seasons. Surely, we must incorporate these measures of variability to understand the systems properties that represent wider levels of organization.

Fortunately, the laws of statistics free us from measuring everything, and we can work with samples, even if that means learning to manage uncertainty. It is curious that people are enraptured by the filigree of the smallest nature beings, like those that Haeckel, one of the patriarchs of ecology, drew more than a century ago, while we marvel watching a sublime landscape in which we barely distinguish the living organisms. Both visions are intimately fitted, not like Russian dolls that identical in their form, but like a tapestry full of knots that are continuously made and undone.
. Eduardo Milla CCBY")

