Why do we want so many different beetles?


Evolution leads to an increasing number of species, and that's why it is so difficult for us to know how many of them inhabit the Earth. Should we spend efforts to conserve all of them or would it be enough with just a few?
![]()
To Jordi and Olga.
In 1992 Edward O. Wilson popularized the concept of 'biodiversity' thanks to his fantastic book The Diversity of Life. Since then, the conservation of species has been incorporated into the values of our society. But what is the magnitude of that effort? Knowing what is the number of species that inhabit the planet can help us answer the question. Our society is attracted to numbers, and the more extreme they are, the greater the attraction they inspire. In 2015, TW Crowther and collaborators obtained a great success in the media with an article published in Nature in which they mapped the density of trees on Earth and estimated at 3.04 · 1012 the total number of living trees throughout the planet. Quickly we had to calculate the number of trees in our Spanish forests—obtaining their corresponding popularity quota. For those interested in the numbers we will say that this value was approximately 7 · 109.
But these estimates have limitations, since they are based on a series of assumptions that are not always fulfilled. Jonathan Chase recently presented a paper in a conference about Biogeography in which he discussed the problems of extrapolations that calculate the number of species that live on Earth. These extrapolations have a lot of uncertainty and values oscillate between 3 and 100 million species—one of the recent calculations in 2011 gave a figure of 8.7 million eukaryotes.
To obtain these numbers a common method is to extrapolate to larger areas, and ultimately the entire planet, the number of species —what we call richness—identified in a known area. This method allows us to have an idea of the number of species that have not yet been identified, basically because the sites in which they live have not yet been prospected. This has implications for conservation, because if the habitat that exists in that site is lost, those species also do.
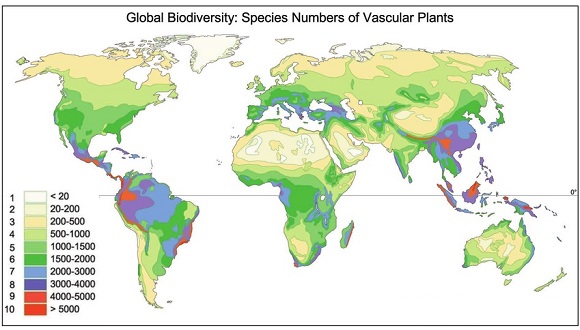
Source: Barthlott, Kier, Rafiqpoor, Kreft, Krüper & Mutke. 2004. Nees Institute for Biodiversity of Plants, University of Bonn. W. Barthlott 1996, 2004. In the paper: Mutke J., Barthlott W. (2005). Patterns of vascular plant diversity at continental to global scale. Biol. Skr. 55:521-531. ISSN: 0366-3612. ISBN 87-7304-304-4.
Logically we must make corrections because these extrapolations are not the same for all groups of living beings. There are groups with many endemic species—such as coleoptera— and therefore prospecting new areas quickly increases their species richness. Other groups—such as mosses— are widely distributed and increasing the area does not increase their number too much. On the other hand, species richness is not the same in all habitats. Therefore, the extrapolation of the number of species to larger areas should analyze separately the relationship between the number of species of each group and the increase of area for each type of habitat, and then weight the sum by the abundance of those habitats. We also have to assess how these extrapolations are affected by habitat fragmentation.
There is also another important consideration: the increase in species richness in larger areas is not linear. It follows a curve with an asymptote that indicates that the environment tends to become saturated with species. Although we prospect with great intensity, it is very difficult to find new species, as naturalists who seek novelties know. The reason is that the number of species in the region that are able to live in that habitat is limited since they have to share the resources among them. This rule is well known and is explained in the first classes of general ecology courses. It is quite curious that in a specialized conference the speaker had to remind listeners of basic ecological principles, supported by more or less sophisticated mathematical models and large databases. I suppose it indicates that there are principles robust enough to persist as science progresses, although new, bright methods momentarily condemn them to oblivion.
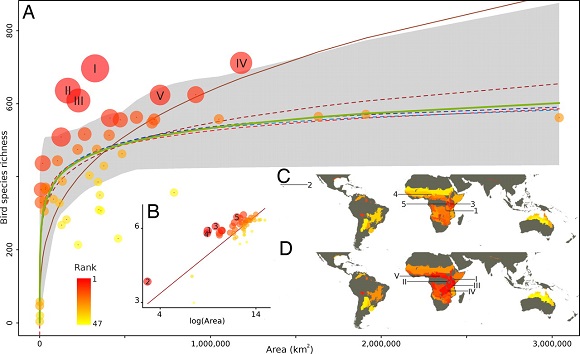
Source: Guilhaumon F., Gimenez O., Gaston KJ., Mouillor D. (2008). Taxonomic and regional uncertainty in species-area relationships and the identification of richness hotspots. PNAS 105 (40): 15458-15463. DOI: https://doi.org/10.1073/pnas.0803610105
Jonathan Chase concludes that we are overestimating the number of species that live on Earth. I do not think that we have to worry too much, unless we are really interested in writting that number in the Guinness book of records. The number of species that inhabits the planet is still the highest in the history of the Earth. It is the result of evolution, which tends to generate new lineages provided they are compatible—sometimes even with a high degree of optimization—with the great existing environmental heterogeneity. At the same time, the diversity of the biota tends to increase environmental heterogeneity, generating new habitats or producing new resources. One of the most remarkable facts that has occurred in natural sciences during the last decades is the recognition of a huge number of genotypes that we attribute to different species from their DNA collected in the environment. These molecular methods have been applied to microorganisms, and a multitude of species have been discovered, identified exclusively by their genetic molecules—technically known as OTUs - Operational Taxonomic Units. Although these methods are not without shortcomings, in general scientists recognized that the number of microbial species is greater than expected. For instance, it has been estimated that in a liter of seawater there may be 20,000 of these "molecular species" and hundreds of them have even been found in rocks at depths of 1,000 meters or more. Currently there are estimates of the global biodiversity of the Earth that reach values between 1 · 109 and 6 · 109 species.
In any case, the conservation of biodiversity has enough work even with the species that we know. Although the number of unknown species was not really as huge as the estimates say, there is no doubt that our knowledge of biodiversity is still incomplete. We must work with that premise and with the certainty that time runs against us. The ignorance of the detail of the biology of many species—of the unknown ones, but also of many of those described by naturalists—, together with the evidence that biodiversity has an important role in the regulation of the functioning of the ecosystems has promoted the success of the concept of functional species—or type of functional species. They are species grouped by those of their characteristics that have a similar implication in ecosystem functioning. This approach has remarkably succeed among researchers who intend to quantify the functioning of the biosphere on a global scale. For example, when calculating Carbon balances, it is essential to determine the primary production of ecosystems, which is basically due to the photosynthetic assimilation of atmospheric CO2 by vegetation. From this point of view, all species that photosynthesize could be put together in a large functional group.
Obviously, this approach can cause horror to a naturalist in love with biodiversity and the particularities of evolutionary responses. But for someone who wants to measure the primary production of all the forests in the world, it is not so relevant if the organism carrying out the photosynthesis is a pine or an oak. In fact, the models that describe the functioning of ecosystems on a global scale have progressed a lot and are considering functional subgroups with differentiated physiological behavior.

Source: Friends of the Earth International (CC BY 2.0)
Soon the question arose about the importance of species diversity when determining the ecosystem flows, such as primary production. If different species do similar functions, a few of them would be enough for ecosystems to work. It is what was called functional redundancy. In addition, it is quite common that the main species responsible for a particular flow of energy or matter are abundant. For example, in crops that need pollinators, only a small number of dominant insect species are needed to obtain a good crop. This rationale made nervous to those who fought to preserve the biodiversity of the planet. Is it worth all the effort to conserve such a great variety of species when, for all practical purposes, they do the same, assuming they really do something? Obviously, the approach contains many simplifications, but it brings important concerns. The first simplification is to assume that there is a unique ecosystem function that deserves to be preserved. Obviously it is not true, and redundant species for a function do not have to be redundant for another one. In addition, the level of species redundancy does not have to be the same for all functions. There are some functions that are performed by a few species, such as the fixation of inert nitrogen from the atmosphere to suitable forms for plants. Only, a small number of plants species intervene in this process, in combination with some soil microbes. If they disappear, the consequences are disproportionate. They are the so-called key species.
The approach corresponds to one of the most attractive research lines of the last decades: to know the contribution of biodiversity in the functioning of ecosystems. It was based on theoretical ideas that advocated a positive relationship between biodiversity and the functioning of ecosystems. The reasoning was based on the complementarity of the species: what one species could not do, another could do. Edward O. Wilson himself—along with his colleague Robert MacArthur, with whom he developed simple and successful mathematical models to explain species richness in insular environments—had supported this idea. Paul Ehrlich and his wife Anne also introduced the idea that redundancy had an imprecise limit, like the screws that hold the fuselage of an airplane: we can lose some pieces and the plane continues to fly, until the fall of a single additional screw can lead to the catastrophe suddenly and at any moment. Nor was an idiosyncratic behavior of each ecosystem discarded, with the consequent difficulties to establish general rules that would help to establish a priori conservation strategies. However, the existence of functional redundancy between species was there.
Any critical ecologist wonders how relevant it is to the ecosystem functioning, not to mention to the biosphere as a whole, that a ladybug has seven or eight black dots. Multiple experiments have been conducted manipulating the number and type of species in field, greenhouse or laboratory conditions—the so-called mesocosms—and measuring the functional properties of the ecosystem. Large field databases, such as forest inventories, with information on biodiversity and primary production measures have been analyzed. Theoretical models and numerical simulations have been developed. Although there are many details to be learned about the involved mechanisms, there is a consensus that the contribution of biodiversity to the functioning of ecosystems is saturated over a certain number of species. That is, the hypotheses of the positive relationship and that of redundancy are not mutually exclusive and both work at to some extent.

Author: Francisco Lloret
In reality, redundancy is not a problem for the conservation of biodiversity, but quite the opposite. Redundancy allows maintaining the functioning of ecosystems, one of whose main qualities is the regulation of the flows of matter and energy over time, since the conditions of the environment change both in space and time. The redundancy of biodiversity provides insurance against an inexorably changing environment. A species that is redundant at one time can become a key species under different environmental conditions. These new conditions can be detrimental to other species that previously performed similar functions and could even have been dominant. Similarly, a dominant species in one place, and therefore largely responsible for the functioning of the ecosystem, may cease to be so in another nearby location. This is what has been seen in a recent study on the role of pollinators in the production of certain crops on the east coast of North America. Although in a locality, a few species are able to provide enough pollen to obtain a good harvest, when we travel throughout the territory new species are needed to cover that function. If anyone has doubts about how redundancy and key species complement each other to provide stability to ecosystems, I invite you to read the brief text prepared by Paul Ehrlich and Brian Walker in 1998
We return to the classic models that interpreted the essence of life through its diversity, by recognizing the role of combining abundance and rarity. Successful, abundant species represent the optimization of biological solutions in front of concrete conditions. But when the environment changes, and it eventually does, rare species have their chance. It is easy to understand that this same reasoning applies to the role of diversity in human societies. Ramon Margalef explained to us that the interest of biodiversity lies in those rare, latent species that apparently play a limited role in the ecosystem, but that are the biological potential that will provide solutions in future, under different environments. We see that we come back around robust principles—easily understood by all humans—, while science complete them, accumulates evidence and thus expands our capacity to understand the world.


